2023: Volume 3, Issue 3
Past Issues
 Abstract
Abstract  PDF
PDFRoles of Indigenous Arbuscular Mycorrhizal Fungi and Composts in Improving Tomato Tolerance to Water Stress under Controlled Greenhouse Conditions
Meriem Mobaligh1, Mohamed Anli2, Abderrahim Boutasknit2, Youssef Ait-Rahou2, Redouane Ouhaddou2, Abdelilah Meddich2, Khalid Fares1,*
1Laboratory of Pharmacology, Neurobiology, Anthropobiology and Environment, Faculty of Sciences Semlalia, Cadi Ayyad University, Department of biology, Marrakech 40000, Morocco
2Laboratory of Agro-Foods, Biotechnologies and Valorisation of Plants Bioresources, Department of Biology, Faculty of Sciences Semlalia, Cadi Ayyad University, Department of biology, Marrakech 40000, Morocco
*Corresponding Author: Khalid Fares, Laboratory of Pharmacology, Neurobiology, Anthropobiology and Environment, Faculty of Sciences Semlalia, Cadi Ayyad University, Department of biology, Marrakech 40000, Morocco; Tel: +212-661-232-285; Email: [email protected]
Received Date: October 11, 2023
Publication Date: October 19, 2023
Citation: Mobaligh M, et al. (2023). Roles of Indigenous Arbuscular Mycorrhizal Fungi and Composts in Improving Tomato Tolerance to Water Stress under Controlled Greenhouse Conditions. Catalysis Research. 3(3):15.
Copyright: Mobaligh M, et al. © (2023).
ABSTRACT
The objective of the present study was to demonstrate the influence of natural fertilizers based on arbuscular mycorrhizal fungi (AMF) and composts as well as chemical fertilizer (NPK) on growth and tolerance of tomato seedlings to water stress. A greenhouse experiment was conducted with tomato seedlings, which were grown in the presence of AMF and/or composts and NPK alone under two water regimes: favorable water regime (75% field capacity (FC) and water stress (35% FC).
The results obtained water stress negatively affected the tomato growth, physiology and biochemistry. However, AMF inoculation combined with NPK or composts improved the total dry biomass of tomato seedlings under normal and drought stress conditions compared to the respective controls. In addition, tomato plants physiology was significantly increased when AMF was combined with C4 or C5 by improving the photosynthetic pigments content (385.2% and 337.4%) as well stomatal conductance (48.3 % and 54.3 %) and chlorophyll fluorescence (43.5 % and 17.2 %) under drought stress conditions compared to the control. Control plants were significantly the most affected by water stress resulted in increased hydrogen peroxide (H2O2) and malondialdehyde (MDA) content. The plants treated with composts as well as AMF alone or combined with composts showed a better capacity of tolerance of water stress by a significant accumulation of soluble sugars, proteins and a high activity of antioxidant enzymes, acting as a control system of reactive oxygen species and thus providing protection against oxidative stress under drought stress. These results highlight the importance of using these biofertilizers, especially in bipartite combination of composts and AMF in improving tomato adaptation to water stress.
Keywords: Water stress, arbuscular mycorrhizal fungi, compost, physiological and biochemical parameters, antioxidant enzymes
INTRODUCTION
Currently, the whole world suffers from agricultural soil degradation and the decrease of its fertility due to intensive agriculture, extreme use of chemical fertilizers as well as pesticides, deforestation and inorganic waste discharges (Kouba et al., 2018; Ouachoua and Karkouri, 2020) [1,2]. Without forgetting the intervention of climate change in exacerbating the situation (Ait-El-Mokhtar et al., 2022a; Borjas-Ventura et al., 2020) [3,4]. The main problems accompanied by the decrease in soil fertility are the loss of soil organic matter, drought and salinity, which could hinder plant growth and development by increasing their susceptibility to nutritional imbalances and diseases (Chiappero et al., 2019; Zhang et al., 2020) [5,6]. Drought stress is a severe environmental challenge for sustainable agricultural production under the present climate change conditions (Delshadi et al., 2017) [7]. Water deficiency disrupts plant growth and development and causes physiological disturbances such as lack of water, stomatal closure, and solute accumulation inside the cell that negatively impact the plant biological system's functioning (Begum et al., 2020; Liu et al., 2021; Saleem et al., 2022) [8-10]. The water deficit adaptation mechanisms put in place by the plant are limited in the case of a prolonged water deficit, and the adverse effects of water deficit cause soil poverty in mineral nutrients and organic matter and consequently crop productivity reduction.
To fulfill demand for agricultural crops, industrialized countries' farming activities have experienced significant changes. On the one hand, high rates of synthetic fertilizers application, as well as pesticides, irrigation, and short crop rotations, have enhanced yields and contributed to the reduction of hunger in these countries (Debaeke et al., 2017; Tilman et al., 2002) [11,12]. Modern high-input agriculture, on the other hand, has resulted in a slew of environmental issues. Groundwater pollution and eutrophication of aquatic systems as a result of soil erosion, nutrient leaching, and run-off from agricultural fields are major concerns in this context (Thonar et al., 2017) [13]. Furthermore, because fertilizer manufacture and use generate large greenhouse gas emissions, the widespread use of synthetic fertilizers contributes to climate change. Furthermore, natural fixation mechanisms in soil reduce the effectiveness of some plant nutrients, such as phosphorus (P), as only a percentage of the given P fertilizer is taken up by plants (Ait-El-Mokhtar et al., 2022c) [14]. A significant amount of P fertilizers is adsorbed to iron, aluminum oxides, or calcium in the soil, depending on the pH and mineralogy, and is no longer immediately available to the plant (Mahmood et al., 2022) [15]. As a result, additional fertilizer inputs are frequently kept on available to suit the crop's needs. Finally, P fertilizers, which are widely employed in modern agriculture, are considered nonrenewable resources having a finite supply, with reserves concentrated in a few nations throughout the world (Ait-El-Mokhtar et al., 2022c; Reid et al., 2021) [14,16]. This situation would be aggravated in the coming years due to climate change. As a result, effective recycling and prudent resource use are essential to promote agricultural sustainability.
For this, it is crucial to assess the effectiveness of using organic amendments capable of mitigating the severity of crop damage caused by abiotic stress including drought stress and ensure the world population food security (Boutasknit et al., 2021; Ullah et al., 2021) [17,18]. Among the current new technologies in agriculture, there is the application of biostimulants based on organic amendments and symbiotic microorganisms (Cozzolino et al., 2021; Vahedi et al., 2021) [19,20]. Soil amendment with composts is a biological way to combat the negative impacts of water deficit on plant growth (Han-Song et al., 2010; Mata-González et al., 2002; Schütz et al., 2018) [21-23]. Indeed, organic amendments, through their composition in micro, macro-nutrients and organic matter, intensify the plants defense system by allowing them to maintain their main functions (Anli et al., 2020; Boutasknit et al., 2020a; Chaichi et al., 2018) [24-26]. In addition, the use of organic amendments allows plants to better maintain their water content and stimulate their enzymatic and non-enzymatic antioxidant system, which play a key role in plant tolerance to different stress conditions (Abd–Elrahman et al., 2022; Ait-El-Mokhtar et al., 2022b; Elfadil et al., 2022) [27-29]. In recent decades, the use of symbiotic microorganisms has become a common practice in areas suffering from drought and is considered both an economically and ecologically beneficial strategy for the regeneration and restoration of soil fertility and productivity (Boutasknit et al., 2020b; Cozzolino et al., 2021) [30,31]. In addition, the contribution of symbiotic microorganisms such as arbuscular mycorrhizal fungi (AMF) can have a potential effect on plant growth as well as stress tolerance through the increase of photosynthetic rate, nutrient and water uptake, as well as physicochemical and biological properties of the soil (Chouyia et al., 2022; Lahbouki et al., 2022) [32,33]. Many recent studies have focused on exploiting arbuscular mycorrhizal fungi (AMF) to bio-protect plants, mitigate the negative effects of drought and ensure soil fertility and productivity (Zou et al., 2015) [34].
Therefore, the present work aims to evaluate the impact of different composts, NPK fertilizer and their combination with arbuscular mycorrhizal fungi (AMF) on the development and tolerance of tomato to water stress under greenhouse conditions.
MATERIAL AND METHODS
Plant Material and Applied Treatments
For surface-sterilization, tomato seeds variety Campbell 33 (Lycopersicon esculentum) were immersed in 5% sodium hypochlorite for 10 min, followed by five washings with sterilized distilled water. They were then placed in Petri dishes containing moistened paper and incubated at 28 °C for 5 days in obscurity for 5 days. The germinated seeds were then placed in trays containing autoclaved peat during 2 weeks. Tomato seedlings of two leaf stage were transferred to plastic pots of 5 kg of previously sterilized soil (for 3 h at 180 °C), which had the following characteristics: electrical conductivity (EC): 190 µS cm−1, pH: 8.60, Total organic carbon: 1.85%, available phosphorus: 11 mg/kg. Different composts used were prepared according to Mobaligh et al. (2021) [35]. The physicochemical characteristics of the five composts tested in this study were given in Table 1.
Table 1: Physicochemical characteristics of the used composts.
|
Parameters |
C1 |
C2 |
C3 |
C4 |
C5 |
|
Humidity (%) |
53.28 ± 0.31 |
45.46 ± 0.14 |
42.15 ± 0.16 |
40.67 ± 0.23 |
40.50 ± 0.32 |
|
pH |
8.60 ± 0.01 |
8.79 ± 0.01 |
8.82 ± 0.02 |
8.90 ± 0.01 |
8.90 ± 0.02 |
|
Total organic matter (% DW) |
52.99 ± 0.49 |
37.96 ± 0.72 |
31.87 ± 0.09 |
24.57 ± 0.11 |
25.04 ± 0.43 |
|
Total organic carbon (% DW) |
29.44 ± 0.32 |
21.09 ± 0.15 |
17.70 ± 0.96 |
13.65 ± 1.15 |
13.91 ± 0.08 |
|
Available phosphorus (mg/kg DW) |
175.99 ± 0.83 |
139.73 ± 1.30 |
118.52 ± 0.72 |
105.75 ± 2.12 |
111.03 ± 1.66 |
|
Ni (mg /kg DW) |
6.31 ± 0.82 |
2.90 ± 0.20 |
7.28 ± 0.16 |
8.98 ± 0.35 |
9.25 ± 1.25 |
|
Cu (mg /kg DW) |
26.19 ± 1.00 |
12.90 ± 0.10 |
17.74 ± 0.14 |
16.8 ± 0.32 |
16.45 ± 0.57 |
|
Pb (mg /kg DW) |
1.45 ± 0.15 |
0.90 ± 0.04 |
0.67 ± 0.03 |
0.66 ± 0.02 |
0.84 ± 0.04 |
|
Zn (mg /kg DW) |
95.92 ± 5.48 |
50.50 ± 1.60 |
125.63 ± 1.59 |
132.69 ± 2.39 |
121.56 ± 10.53 |
|
Cr (mg /kg DW) |
5.44 ± 1.92 |
2.20 ± 0.10 |
10.75 ± 1.79 |
14.89 ± 1.26 |
13.76 ± 0.83 |
|
As (mg /kg DW) |
8.89 ± 0.02 |
5.30 ± 0.01 |
7.68 ± 0.06 |
7.29 ± 0.04 |
6.57 ± 0.29 |
|
Ca (% DW) |
1.67 ± 0.00 |
21.10 ± 0.00 |
2.23 ± 0.00 |
16.98 ± 0.00 |
17.78 ± 0.00 |
|
K (% DW) |
5.18 ± 0.00 |
5.66 ± 0.00 |
2.94 ± 0.00 |
4.84 ± 0.00 |
4.25 ± 0.00 |
|
Mg (% DW) |
0.57 ± 0.00 |
1.40 ± 0.00 |
1.16 ± 0.00 |
2.80 ± 0.00 |
2.52 ± 0.00 |
|
Na (% DW) |
5.38 ± 0.00 |
3.21 ± 0.00 |
5.10 ± 0.00 |
1.84 ± 0.00 |
1.95 ± 0.00 |
|
Fe (% DW) |
0.00 |
0.00 |
0.22 ± 0.00 |
0.31 ± 0.00 |
0.29 ± 0.00 |
DW: dry weight; C1: green waste + leachate from landfill; C2: green waste + leachate from landfill + sugar lime sludge (20%); C3: green waste + leachate from landfill + phosphate washing sludge (20%); C4: green waste + leachate from landfill + phosphate washing sludge (50%) and C5: green waste + leachate from landfill + phosphate washing sludge (50%) + olive mill wastewater.
The study used a randomized completely block design with two variables: biofertilizer treatments and water stress levels. During transplanting, compost treatments, soil was mixed and homogenized with compost at a rate of 5%. The AMF treatments were inoculated with 20 g of fresh inoculum from the native mycorrhizal consortium containing root fragments and spores as close to the roots as possible during transplanting, and the chemical fertilizer treatment received 7 g of NPK per pot. The plants were grown in the greenhouse of the Faculty of Science Semlalia, Cadi Ayyad University, Marrakesh, Morocco, for four months under controlled conditions. Two watering conditions were used: well-watered (75% field capacity (FC) and water stress (35% FC). 14 treatments were applied with five replicates for each treatments (Table 2).
Table 2: Treatments received by the two water regimes used in the experiment.
|
75% field capacity |
||||||||||||||
|
Treatments |
-AMF |
+AMF |
||||||||||||
|
Control |
NPK |
C1 |
C2 |
C3 |
C4 |
C5 |
Control |
NPK |
C1 |
C2 |
C3 |
C4 |
C5 |
|
|
35% field capacity |
||||||||||||||
|
Treatments |
-AMF |
+AMF |
||||||||||||
|
Control |
NPK |
C1 |
C2 |
C3 |
C4 |
C5 |
Control |
NPK |
C1 |
C2 |
C3 |
C4 |
C5 |
|
Control: not inoculated and not amended; NPK: amended by chemical fertilizer; C1: (Green waste + Leachate); C2: (Green waste + Leachate + sugar lime sludge (20%)); C3: (Green waste + Leachate + Phosphate sludge (20%)); C4: (Green waste + Leachate + Phosphate sludge (50%)); C5: (Green waste + Leachate + Phosphate sludge + oil wastewater (5 L)); AMF: arbuscular mycorrhizal fungi consortium.
Mycorrhizal colonization measurement
Fresh roots were cut into 1 cm segments, rinsed, and cleaned in 10% KOH for 30 min at 90 °C. The segments were acidified for 10 min with 5% lactic acid, dyed for 30 minutes at 90 °C with 0.05% (w/v) Trypan blue, and then microscopically examined for root mycorrhizal colonization. In 20 randomly chosen root segments (1 cm length) per glass slide, the frequency of fungal structures in the root system (F%) and the intensity of mycorrhizal colonization (M%) were examined. According to McGonigle et al. [36] (1990), mycorrhizal parameters (F% and M%) were computed from root segments as follows:
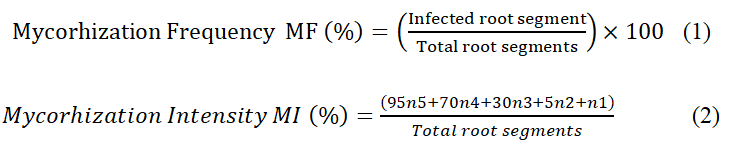
where n is the number of fragments assigned with the index 0, 1, 2, 3, 4, or 5, with the following infection rates: 100 > n5 > 90; 90 > n4 > 50; 50 > n3 > 10; 10 > n2 > 1; 1 > n1 > 0.
Evaluation of plant growth
The tomato seedlings were harvested four months after transplantation. The root system was separated from the shoots part and biometric data (plant height, root elongation and dry weight of shoot and root parts) were determined.
Stomatal conductance and photosynthetic efficiency evaluation
Before the harvest, stomatal conductance (gs) was measured with a porometer (Leaf Porometer LP1989, Decagon Device, Inc., Washington, USA) on five replicates per treatment on a sunny day between 09:30 and 11:00 am as described by Harley et al. (1992). With five repetitions per treatment, photosynthetic efficiency was measured on the third fully developed leaves using a portable fluorometer (Opti-sciences OSI 30p). Transmission at 650 nm on a leaf area of 12.5 mm² was used to determine this parameter. The quantum yield (Fv/Fm = (Fm - F0)/Fm) were represented by the Fv/Fm values, where Fm and F0 are the maximum and initial quantum yields of dark-adapted leaves, respectively (Hosseinzadeh et al., 2015) [37].
Photosynthetic pigments content determination
The extraction of photosynthetic pigment was carried out according to Arnon (1949) [38] method. Leaf samples were ground in 80% acetone and the extract centrifuged for 10 minutes at 10,000 g. A spectrophotometer was used to measure the absorbance of the supernatant at three wavelengths: 480, 645, and 663 nm. Based on the observed absorbance values, the photosynthetic pigment content was determined. As a blank, 80% acetone was utilized.
Hydrogen peroxide content and lipid peroxidation evaluation
The content of hydrogen peroxide (H2O2) in leaves was determined spectrophotometrically using the Velikova et al. (2000) method. Leaf material (0.25 g) was homogenized in 5 mL of 10% (w/v) trichloroacetic acid (TCA) before being centrifuged for 15 minutes at 15,000 g. The hydrogen peroxide concentration of the recovered supernatant was determined. 1 mL potassium iodide (1 M) and 0.5 mL potassium phosphate buffer (10 mM, pH 7) were added to the supernatant. The absorbance was measured at 390 nm after a 1 h of incubation in the dark. As a control, 10% TCA was used. The content of malondialdehyde (MDA) was utilized to determine lipid peroxidation using the Dhindsa et al. (1981) [39] method. Leaf samples (0.25 g) were crushed in 10 mL of 0.1 % TCA and centrifuged for 10 minutes at 4 °C at 18,000 g. The supernatant (2 mL) was combined with 2 mL of 20% TCA solution containing 0.5% thiobarbituric acid (TBA). The mixture was boiled for 30 min at 100 °C, quickly cooled, and then cleared by centrifugation at 10,000 g for 10 min. The MDA concentration was determined by measuring the absorbance of the supernatant at 532 and 600 nm.
Soluble sugars content evaluation
The leaf material (0.1 g) was ground in 4 mL of 80 % ethanol and centrifuged for 10 minutes at 5000 rpm. The pellet was re-suspended in 2 mL of ethanol and centrifuged after collecting the supernatant. Using the Dubois et al. (1956) [40] approach, supernatants were used to measure soluble sugars. 5 mL of concentrated sulfuric acid and 1 mL of 5% phenol solution were mixed with 1 mL of supernatant. A spectrophotometer was used to measure the absorbance after 5 minutes at 485 nm (UV-3100PC). As a standard, glucose was used.
Measurement of proteins content
Bradford's (1976) [41] technique was used to determine the soluble protein content. Plant samples (1 g) were homogenized in 4 mL of 1 M phosphate buffer (pH 7.2) and centrifuged for 15 minutes at 4 °C at 18,000 x g. Spectrophotometer cuvettes were pipetted with supernatants and dyes, and absorbance was measured at 595 nm.
Enzymatic antioxidant activities evaluation
The peroxidase (POX) activity was determined using Polle et al. (1994) [42] technique. In 0.1 mL enzyme extract, 3 mL 1 M phosphate buffer (pH 7.0), 20 mM guaiacol, and 40 mM H2O2 were used in the reaction mixture. The capacity of POX to convert guaiacol to tetraguaiacol was measured at 470 nm. An absorbance change of 0.01 unit min-1 was considered as one unit of POX activity.
The Hori et al. (1997) [43] technique was used to assess polyphenoloxidase (PPO) activity at 410 nm while monitoring catechol oxidation. 600 µL of 0.1 M catechol and 100 µL enzyme extract were added to 3 mL of potassium phosphate buffer (0.1 M, pH 7.0). The activity of the enzyme was determined in µmol of catechol min-1 mg-1 of protein.
The activity of catalase (CAT) was assessed using Aebi (1984) [44] method. CAT activity was determined as an absorbance decrease caused by H2O2 decomposition for 3 min at 240 nm. The reaction solution consisted of 100 μL of vegetal extract, 0.1 mM potassium phosphate buffer (pH 7.0), 20 mM H2O2, and 0.1 mM EDTA.
Soil Analyses
Soil physicochemical parameters were examined on samples taken near the roots during harvest. A pH meter and an electrical conductivity (EC) meter were used to measure the pH and electrical conductivity (EC) of a diluted soil suspension of 1/10 (v/v). Total organic carbon (TOC) and organic matter (OM) were determined using the method of Nelson and Sommers (1996) [45], and available phosphorus (AP) was determined using the technique of Olsen and Sommers (1982) [46].
Statistical Analysis
All data were examined using the ANOVA technique and the Tukey's Honestly Significant Difference (HSD) test with CO-STAT Statistical Software (CoStat 6.4) at a 5% significance level. To check whether the residuals were normal, the Shapiro-Wilk test was performed. XLSTAT v. 2019 was used to perform the principal component analysis (PCA).
RESULTS
Mycorrhization Parameters Obtained results revealed that water stress negatively affects tomato roots colonization by AMF. However, no significant different was shown in all treatments received AMF inoculation (Figure 1).
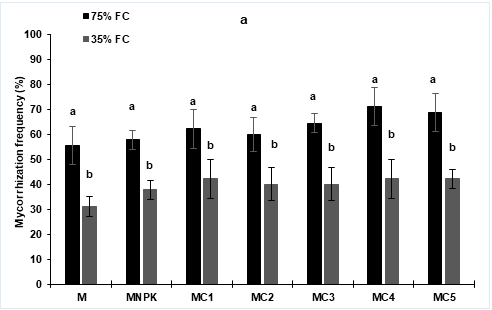

Figure 1: Mycorrhization frequency (a) and intensity (b) of tomato (Lycopersicon esculentum) seedlings after 4 months of composts and NPK application and AMF inoculation under two water regimes (75 and 35% FC). Tukey’s HSD test at P < 0.05 was used to compare mean values ± SD (Standard Deviation) to each other.
GROWTH TRAITS
Results of measured growth parameters were presented in Figure 2. The shoot height was significantly improved under drought stress conditions in plants treated by MC4 compared to the control and NPK (Figure 2a). In addition, under the same conditions, root elongation showed no significant difference for all applied treatments compared to the control (Figure 2b). In addition, total dry weight was significantly reduced under drought stress conditions. However, applied treatments, especially AMF combined with NPK or composts showed significant improvement of total dry weight under drought stress conditions compared to the control. The highest improvement was recorded in tomato plants treated by MC1 (30%) compared to the control under water deficit conditions (Figure 2c).

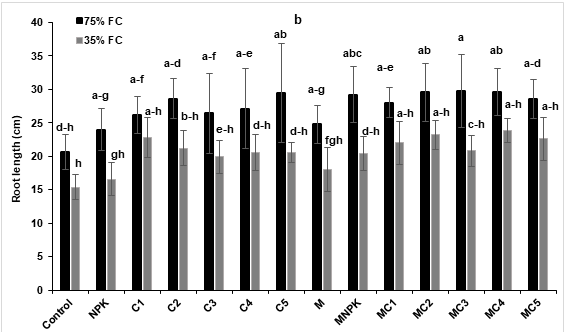
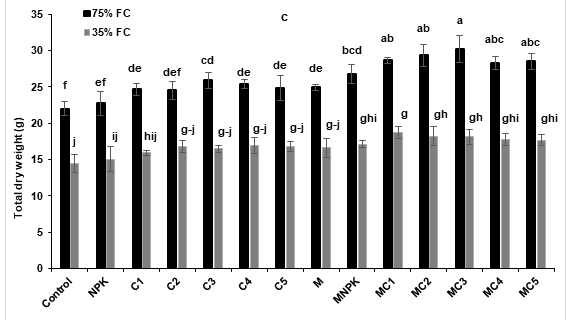
Figure 2: Effects of composts and NPK application and AMF inoculation on tomato (Lycopersicon esculentum) growth parameters: (a) shoot height (b) root length and (c) total dry weight under two water regimes (75 and 35% FC). Tukey’s HSD test at P < 0.05 was used to compare mean values ± SD to each other.
Physiological traits
Obtained results showed that water stress significantly decreases stomatal conductance and chlorophyll fluorescence (Figure 3). However, tomato seedlings treated by NKP, C3, MC2, MC3, MC4 and MC5 revealed a significant improvement of this traits compared to the control under stressed conditions (Figure 3a). Additionally, chlorophyll fluorescence was increased in plants treated by C1, C2, M, MC1, MC2, MC3, MC4 and MC5 compared to the control and NPK treatments (Figure 3a).
The results presented in Fig. 4 showed that water stress negatively affects photosynthetic pigments namely chlorophyll a and b, total chlorophyll and carotenoid contents. The application of AMF and composts, especially MC2, MC3, MC4 and MC5 treatments, showed significant improvement of chlorophyll as compared to the control and NPK treatments, respectively (Figure 4a). Moreover, chlorophyll a and total chlorophyll content (T Chl) were significantly improved by C4, C5, MC1, MC2, MC3, MC4 and MC5 treatments under water-limiting (35% FC) conditions compared to the control and NPK (Fig. 4b and c). As for carotenoid content, it was increased by all treatments compared to the control and NPK treatments except M and MNPK treatments, respectively (Figure 4d).

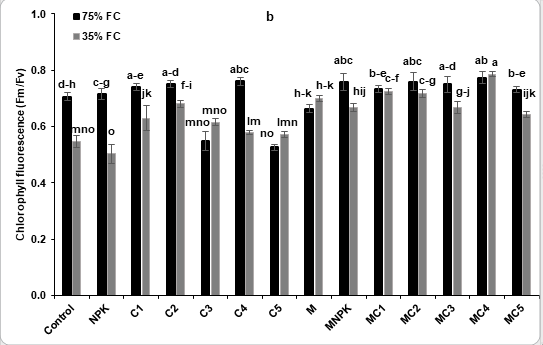
Figure 3: Effects of compost and NPK application and AMF inoculation on physiological (a) stomatal conductance and (b) chlorophyll fluorescence of tomato (Lycopersicon esculentum) seedlings under two water regimes (75 and 35% FC). Tukey’s HSD test at P < 0.05 was used to compare mean values ± SD to each other.
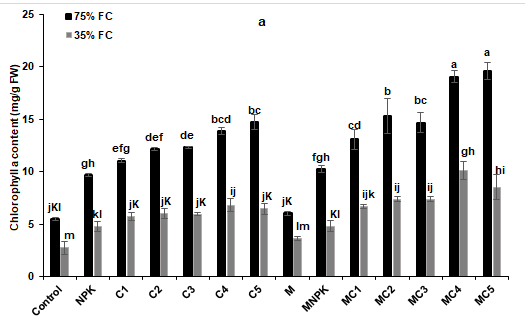
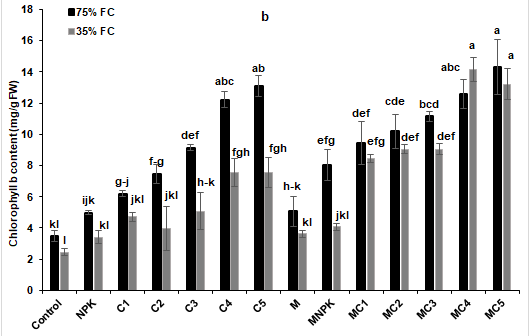
Figure 4: (a) chlorophyll a, (b) chlorophyll b, (c) total chlorophyll, and (d) carotenoid content in leaves of tomato plants under two water regimes (75% and 35% FC) and further grown without or with composts, NPK and arbuscular mycorrhizal fungi. Tukey’s HSD test at P < 0.05 was used to compare mean values ± SD to each other.
Biochemical Traits
The defense of tomato plants against water stress is not only limited by physiological responses but rather by a biochemical response that translates into an increase in proteins and sugars. According to the results in Figure 5, this response differs from one treatment to another. Under the dry soil conditions (35% FC), the protein and sugar contents were negatively affected. On the other hand, this effect was overcome by the different applied treatments than the control and NPK treatments except the M and MNPK treatments for protein and M treatment for sugar, respectively (Figure 5a and b). The highest improvement was recorded in MC4 and MC5-plants by 401 and 372% for proteins and 163 and 172% for sugars, respectively, compared to the control under drought stress conditions.
The evaluation of antioxidant enzymes was estimated to determine to what extent tomato plants will contract the deleterious effect of water stress. Catalase (CAT), peroxidase (POX) and polyphenol oxidase (PPO) were estimated under both conditions (75% and 35% FC). The significant increase in the activity of these three enzymes was pronounced in control plants as well as in tomato plants treated with M alone under stressed conditions (Figure 6).
In order to know the deleterious effects caused by water stress on tomato plants, analyses of the two stress markers were considered by evaluating H2O2 and MDA contents (Figure 7). An increase of these stress markers was noticed following the exposure of tomato plants to severe water stress (35% FC), especially in control and NPK plants for H2O2 (Figure 7a) and control plants for MDA (Figure 7b), respectively compared to the other treatments. However, biostimulants application was able to reduce these contents and the highest reduction was recorded in plants treated with MC5 combination by 25% for H2O2 and 50% for MDA, respectively, compared to the control under drought stress conditions.


Figure 5: (a) total soluble proteins and (b) sugars content in tomato plants under two water regimes (75 and 35% FC) after application of composts, NPK and arbuscular mycorrhizal fungi. Tukey’s HSD test at P < 0.05 was used to compare mean values ± SD to each other.

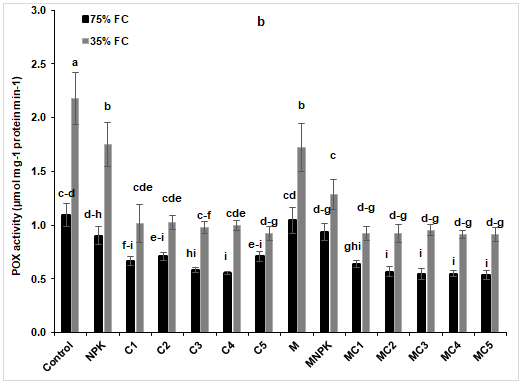

Figure 6: (a) catalase, (b) peroxidase (POX) and (c) polyphenol oxidase (PPO) activities in tomato plants under two water regimes (75 and 35% FC) after application of composts, NPK and arbuscular mycorrhizal fungi. Tukey’s HSD test at P < 0.05 was used to compare mean values ± SD to each other.
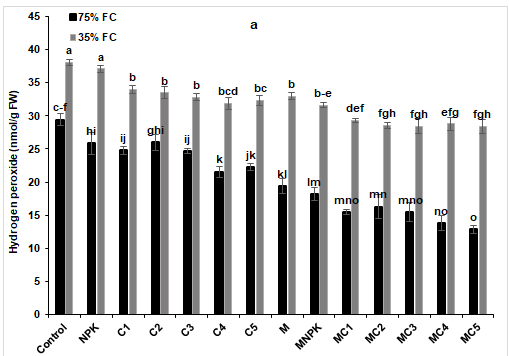
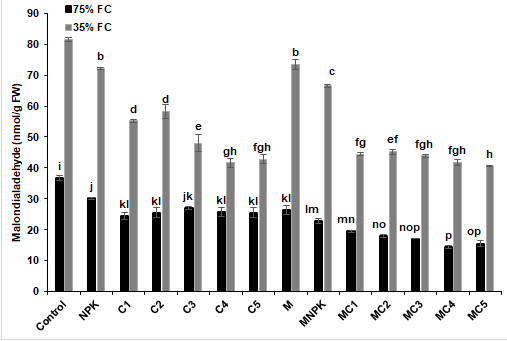
Figure 7: (a) Malondialdehyde (MDA) and (b) hydrogen peroxide (H2O2) content in in tomato leaves under two water regimes (75 and 35% FC) after application of composts, NPK and arbuscular mycorrhizal fungi alone or in combination. Tukey’s HSD test at P < 0.05 was used to compare mean values ± SD to each other.
Effect of composts and NPK application and AMF inoculation on soil characteristics
The findings of the soil analysis were shown in Fig. 8 and 9 after four months of composts and NPK application and/or AMF inoculation. The soil pH was significantly increased in soil treated by NPK fertilizer in both well-watered and drought stress conditions, respectively compared to the controls (Figure 8a). In addition, electrical conductivity was significantly improved in soils amended with composts, inoculated with AMF and those treated by composts combined with AMF under normal and stressed conditions than controls (Figure 8b). Moreover, regardless of the water regime, an increase in soil TOC, TOM and P content was observed after 4 months of tomato growing when composts were applied alone or in combination with AMF (Figure 9) (Compared to the control). However, the highest concentrations of P were obtained in soils treated by C4 (1288 and 1540%), C5 (1353 and 1617%), MC4 (1333 and 1627%) and MC5 (1322 and 1643%) under well-watered and drought stress conditions, respectively compared to the controls.

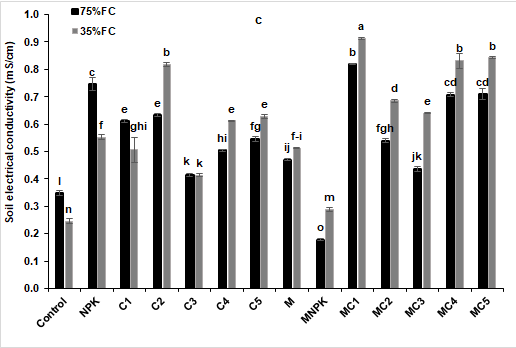
Figure 8: Impact of composts and NPK application and AMF inoculation on soil (a) pH and (b) electrical conductivity at harvest time of tomato grown under two water regimes (75 and 35% FC). Tukey’s HSD test at P < 0.05 was used to compare mean values ± SD to each other.


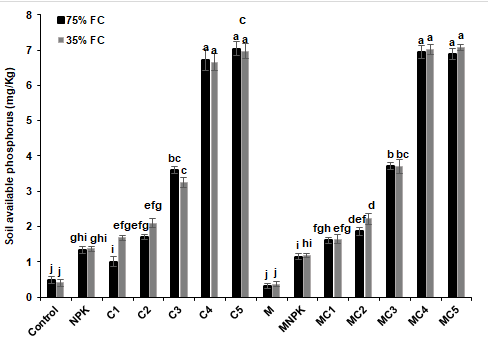
Figure 9: Impact of composts and NPK application and AMF inoculation on soil (a) total organic carbon, (b) total organic matter and (c) available phosphorus content at harvest time of tomato grown under two water regimes (75 and 35% FC). Tukey’s HSD test at P < 0.05 was used to compare mean values ± SD to each other.
DISCUSSION
Drought is one of the most devastating global environmental problems that negatively affect plant growth and development as well as productivity (Khan et al., 2021) [47]. The investigation for mechanisms that can promote plant growth and tolerance to water scarcity at the time of cultivation is therefore critical. Several biological technologies such as the use of biofertilizers (AMF and compost) are currently utilized frequently in agricultural programs to boost plant yield and enhance their resistance to environmental challenges (Köhl et al., 2016; Van Der Heijden et al., 2015) [48,49]. Indeed, symbiosis with AMF increase plant growth and could confer drought resistance due to mycelia proliferating in the soil beyond the rhizosphere, forming a specialized network for acquiring water and mineral nutrients from the soil (Hashem et al., 2019; Meddich et al., 2015) [50,51]. Furthermore, compost contains nutrients that are easily assimilated by the plant such as nitrates, phosphorus, potassium, calcium, and magnesium (Ullah et al., 2021) [18]. It is also able to provide better regulation of oxidative stresses induced by environmental constraints. In light of this, the current study aimed to study the effect of NPK fertilizer, AMF and novel leachate and phosphate sludge composts, alone or in combination, on growth, photosynthesis, metabolite accumulation, alterations in ROS removal mechanisms and soil physicochemical characteristics in tomato in greenhouse trials. Regarding mycorrhization parameters, the native mycorrhizal consortium successfully infected inoculated plants, but no mycorrhizal structures were observed in the roots of uninoculated tomatoes. Root colonization was significantly reduced by application of the 35% FC water regime compared to the 75% FC water regime in the absence or presence of NPK or composts application. This result is in agreement with the finding of Zhang et al. (2018) [52] and Wu et al. (2013) [53] who reported that water stress had a negative effect on root colonization by AMF during the drought period. This decrease in root colonization could be attributed to the inhibition of AMF spores germination caused by the water stress severity (Kilpeläinen et al., 2020) [54]. In addition, the application of NPK and composts showed no difference on root colonization compared to the M treatment alone. This may be explained by the ability of these amendments applied at optimal doses to enhance or maintain root colonization by AMF (Anli et al., 2022; Ziane et al., 2021) [55,56]. This advantageous impact can be attributable to compost's stimulatory effects on AMF biological processes, such as spore germination as well as hyphae spread and and mycelium length (Cozzolino et al., 2016, 2021) [57]. Additionally, according to other studies, the presence of phosphate and humic acid in the compost could promote the development of AMF hyphae (Anli et al., 2021; Pinos et al., 2019) [24,58].
The present study showed that the application of water stress significantly reduced tomato growth. However, all composts used alone or in combination with AMF significantly improved tomato biomass under drought stress conditions compared to the control plants. This is in agreement to the findings of other previous studies that proved the beneficial effect of compost application on the yield and growth of several crops under water stress conditions such as wheat (Verma et al., 2015) [59], quinoa (Benaffari et al., 2022) [60] and date palm (Anli et al., 2020) [61]. Compost constitutes a finely degraded and fully stabilized organic matter that promotes seed germination, growth, flowering and fruiting of a wide range of crops through hormonal effects (Mandal and Pal, 2022) [62]. This enhancement can be attributable to the fact that compost applied as an organic matter source provides enough mineral nutrients for the growth, development and tolerance of plants under abiotic stress (Anli et al., 2020; Olayiwola and Abiodun, 2019) [24,63]. Moreover, AMF could increase the bioavailability of nutrients provided from compost to promote plant growth directly or indirectly from soil to plants via the mycelium structures under drastic environments (Ait-El-Mokhtar et al., 2022b; Duo et al., 2018; Ortuño et al., 2018) [28,52,54]. Furthermore, when compared to the control and chemical fertilizer treatment, the combination of the two amendments with AMF (MC4 and MC5) significantly improved tomato seedling growth, which can be explained by the combined positive effects of composts and AMF in producing a higher allocation of nutrients to plants and their ability to increase phosphorus solubility (Benaffari et al., 2022; Cozzolino et al., 2021) [31,64].
We studied in a comparative way, the effects of AMF, compost and NPK fertilizer on the resistance to water stress of young tomato plants in order to evaluate their adaptation to drought. The best growth improvement was obtained by the combined applications of both MC4 and MC5. If considered separately, composts improved plant growth more, followed by AMF and finally NPK fertilizer. The evaluation of water stress resistance of tomato seedlings showed physiological alteration of tomato plants during the water stress period. The water deficit reduced stomatal conductance and chlorophyll fluorescence. Both parameters were less affected in tomato plants in the presence of biofertilizers under drought stress conditions. These findings were in agreement with those obtained by several researchers (Anli et al., 2020; Boutasknit et al., 2021; Symanczik et al., 2018) [24,65,66]. Stomatal conductance reveals the stomatal opening status and the gas exchange status of the plant. During drought episodes, the plant closes its stomata to avoid damage (Lovisolo et al., 2010) [67]. Our results suggest that plants treated with biofertilizers alone or in combination have an ability to maintain high stomatal conductance and chlorophyll fluorescence under water deficit conditions, which could reflect a better adaptation to this stress through an efficiency of using radiation for photochemical reaction and CO2 assimilation (Bouchemal et al., 2018; Morales et al., 2020; Yang et al., 2020) [68-70]. The leaf chlorophyll and carotenoids were also affected content with the greatest losses observed in control plants during water stress. The biofertilizers used recovered the photosynthetic pigments content of the plants with better improvements recorded in the plants that received the amendments (C4 and C5) in combination with AMF under drought stress conditions. Photosynthetic pigments are very important traits used in several studies to detect the response to water stress in plants (Shafiq et al., 2021; Yang et al., 2020) [71,62]. Water stress limits photosynthetic activity through a lowering of chlorophyll pigment contents. This reduction is either due to a degradation of these pigments, a decrease in their synthesis or to the closure of stomata and the lack of CO2, which are associated with an accentuated production of ROS (Al-Arjani et al., 2020; Yang et al., 2020) [72,63]. The decrease in photosynthetic pigments level under water stress conditions is largely due to damage to chloroplasts by ROS such as O2- and H2O2, which can lead to peroxidation of membrane lipids and consequently, destruction of chlorophyll (Begum et al., 2020; Hasanuzzaman et al., 2020) [8,74], which linked with an enhanced ROS production in the thylakoids (Kiani et al., 2008; Tian et al., 2017) [75,76]. However, our results showed that under water stress conditions, the biofertilizers allowed the plants to still maintain high photosynthetic pigments content compared to the control. The intervention of biofertilizers allowed the tomato plants to restore the photosynthetic system during the period of water stress. This suggests that the use of AMF and compost can protect the photosynthetic system of plants from degradation caused by water stress. Furthermore, significant variations in sugar accumulation were observed under water stress conditions and plants treated by composts alone and AMF+Compost recorded the highest sugar levels for both water regimes (35% FC and 75% FC) compared to the controls. Sugars are considered by several authors as good osmoregulators that may play an important role in osmotic adjustment and plant adaptation to drought damages (Bárzana et al., 2014; Farooq et al., 2020; Shafiq et al., 2021) [71,77,78]. Osmotic adjustment is an early physiological response of plants to water stress that allows cells to remain turgid at very low water potentials through active solute accumulation. They help maintain turgor pressure, which is necessary for cell expansion and synthesis of cell wall components, including cellulose (Thalmann and Santelia, 2017) [79]. According to Sairam et al. (2002) [80], these solutes can interact with cellular macromolecules such as antioxidant enzymes, stabilizing their structure and function.
A drastic increase of H2O2 and MDA was recorded in the leaves during the period of water stress. This is a very usual phenomenon when plants face hostile conditions. Comparing the different treatments applied, the control plants had very high H2O2 and MDA contents than the plants that received AMF and/or composts/NPK, which recorded the lowest values. This indicates that the control plants had experienced a much higher oxidative stress than the plants under different biological treatments. Low H2O2 and MDA accumulation in colonized plants provides evidence for the ability of AMF and/or composts to mitigate oxidative damage caused by ROS and to enhance plant antioxidant defense systems in response to water insufficiency (Benaffari et al., 2022; Boutasknit et al., 2020b; Huang et al., 2014; Ni et al., 2013; Rani et al., 2018; Wu et al., 2013) [30,53,59,80,81,82]. In conjunction with the increase in H2O2 and MDA in the leaves, a decrease in leaf protein under water stress conditions was observed with higher losses in control plants compared to plants with fertilizer treatments. This water stress induced decrease in protein, would be mainly due to ROS damage to proteins (Rani et al., 2018) [82] and an improvement in protein levels by the fertilized plants could be due in part to the ability of the fertilizers used to enhance the antioxidant defense system of the plants that prevents excessive degradation of proteins and other structural plant macromolecules (Boutasknit et al., 2020b; Farooq et al., 2020; Ruiz-Lozano, 2003) [30,70,84]. Our study also showed that under water stress conditions, antioxidant enzyme activities were higher in the control plants than in the plants that received the amendments, especially with the combined C+M application, where the minimum activities of CAT, POX and PPO were recorded. This high antioxidant enzymes activity was proportional to the accumulation of oxidant molecules, especially H2O2, which was higher in the control than in the fertilizer treatments, including AMF and/or composts/NPK treatments. Indeed, to protect themselves against ROS and their intermediates, plant cells at the level of their organelles such as chloroplasts, mitochondria and peroxisomes, develop powerful antioxidant defense systems (Tahiri et al., 2022; Yadav et al., 2010) [85,86]. Previous studies have demonstrated that compost and AMF could play a key role in enhancing the antioxidant system, especially in stressful environments (Boutasknit et al., 2020b; Duo et al., 2018; Hasanuzzaman et al., 2021) [30,64,87]. According to Djébali et al. (2011) [88], the concentration of ROS in plant tissues is the result of a delicate balance between the rates of their production and elimination by the antioxidant system that defines the normal intracellular level of ROS and maintenance at steady state. Instead of having an immediate deleterious effect, this increase in ROS production is likely to be beneficial to the plant if it nevertheless remains under strict control (Bouchemal et al., 2018) [89]. However, under water stress conditions, this balance will be disturbed by the excessive increase of ROS in plant cells. The activation and/or synthesis of antioxidants depends on the species involved, the metabolic state of the plant and the intensity and duration of the stress (Ahmad, 2014; Boutasknit et al., 2021) [66]. This would explain the high activity of antioxidant enzymes (CAT, POX and PPO) coupled with high H2O2 concentrations in the control, due in part to intense stress that disrupted the ROS/antioxidant balance very quickly. Our results suggest that the presence of multiple enzymes performing the same catalytic function is a very important feature that extends the ability to adapt to water stress (Bouchemal et al., 2018; Du et al., 2019) [69,89].
The use of biofertilizers has many advantages for agricultural soils. The present work demonstrated the beneficial value of using AMF and/or compost to improve soil quality without influencing the environment function. In this study, the used soil revealed low TOM and available P. The proposed biofertilizers (composts and AMF) improved the soil physicochemical traits, namely soil TOM and available P under both 75 and 35% FC. These results are probably due to the high organic matter content of composts (Table 1), especially C1 and C2 combined or not with AMF, which showed significant improvement on growth and physiological traits under normal and stressed conditions. This could be also due to the role of AMF in metabolizing various chemical compounds contained in the composts or produced by plant roots like strigolactones and glomalin (Ben-Laouane et al., 2021; Kaushal, 2019; Wang et al., 2021) [17,90,91]. In addition, the present study, application of composts, especially C4 and C5 combined with AMF significantly increased the growth and tolerance of tomato seedlings but also improved the TOM content and P availability in the soil compared to the NPK treatment under drought stress conditions. These obtained results are in agreement with those found in other studies, which reported that plants treated with organic fertilizers combined or not with AMF improved growth and tolerance of plants compared to NPK fertilizer (Kevin et al., 2021; Sun et al., 2018; Yu et al., 2022) [92-94] Composts C4 and C5 with their high richness in humus and mineral nutrients such as P, K, Mg and Ca could also contribute to aggregate stability and improved soil structure and fertility and consequently the plant growth and productivity.
CONCLUSION
The present study allowed highlighting the effectiveness of biofertilizers and NPK input on the growth and adaptation of tomato seedlings to water stress. Based on the results obtained, we can conclude that the fertilizers used, namely AMF, compost and NPK fertilizer significantly improved the growth of tomato plants grown under greenhouse.
The bipartite combination of AMF and composts gave a better growth of the plants compared to the untreated plants or treated with the NPK fertilizer. This concerns all the measured growth parameters. Our results also showed that water stress (35 % FC) is a limiting factor for young growing tomato plants by affecting a large number of physiological and biochemical processes. These young plants were much more vulnerable to a prolonged water deficit if they did not receive fertilizers that can help them adapt to the increased water stress. Application of these fertilizers allowed tomato seedlings to better withstand the water stress and recover physiologically after the water stress period, but significant differences were observed between the applied fertilizers.
The AMF+Compost treatment conferred better resistance to water stress compared to the control plants and the NPK fertilizer treatment, which offers opportunities to exploit these indigenous natural resources for better drought adaptation of tomato plants.
FUNDING
This research received no external funding.
CONFLICT OF INTEREST
The authors declare that they have no conflict of interest.
REFERENCES
- Kouba Y, Gartzia M, El Aich A, Alados CL. (2018). Deserts do not advance, they are created: Land degradation and desertification in semiarid environments in the Middle Atlas, Morocco. J Arid Envir. 158:1–8.
- Ouachoua R, Karkouri J Al. (2020). Application of the environmental vulnerability index ( EVI ) in Ziz- Rheris Watershed , Morocco. Int J Sci Res Multidisci Stud. 6(9):1–9.
- Ait-El-Mokhtar M, Boutasknit A, Ben-Laouane R, Anli M, El Amerany F, Toubali S, et al. (2022a). Vulnerability of oasis agriculture to climate change in Morocco. IGI Global. 1195-1219.
- Borjas-Ventura R, Ferraudo AS, Martínez CA, Gratão PL. (2020). Global warming: Antioxidant responses to deal with drought and elevated temperature in Stylosanthes capitata, a forage legume. J Agro Crop Sci. 206:13–27.
- Chiappero J, Cappellari L, del R, Sosa Alderete LG, Palermo TB, Banchio E. (2019). Plant growth promoting rhizobacteria improve the antioxidant status in Mentha piperita grown under drought stress leading to an enhancement of plant growth and total phenolic content. Industrial Crops and Products. 139.
- Zhang Y, Ding J, Wang H, Su L, Zhao C. (2020). Biochar addition alleviate the negative effects of drought and salinity stress on soybean productivity and water use efficiency. BMC Plant Biol. 20:1–11.
- Delshadi S, Ebrahimi M, Shirmohammadi E. (2017). Effectiveness of plant growth promoting rhizobacteria on Bromus tomentellus Boiss seed germination, growth and nutrients uptake under drought stress. South Afr J Bot. 113:11–18.
- Begum N, Ahanger MA, Zhang L. (2020). AMF inoculation and phosphorus supplementation alleviates drought induced growth and photosynthetic decline in Nicotiana tabacum by up-regulating antioxidant metabolism and osmolyte accumulation. Envir Exp Botany. 176:104088.
- Liu RC, Xiao ZY, Hashem A, Abd_allah EF, Xu YJ, Wu QS. (2021). Unraveling the interaction between arbuscular mycorrhizal fungi and camellia plants. Horticulturae. 7(9):1–12.
- Saleem MH, Wang X, Parveen A, Perveen S, Mehmood S, Fiaz S, et al. (2022). Alleviation of drought stress by root-applied thiourea is related to elevated photosynthetic pigments, osmoprotectants, antioxidant enzymes, and tubers yield and suppressed oxidative stress in potatoes cultivars. Peer J. 10:13121.
- Debaeke P, Pellerin S, Scopel E. (2017). Climate-smart cropping systems for temperate and tropical agriculture: Mitigation, adaptation and trade-offs. Cahiers Agricultures. 26:0–12.
- Tilman D, Cassman KG, Matson PA, Naylor R, Polasky S. (2002). Agricultural sustainability and intensive production practices. Nature. 418:671–677.
- Thonar C, Lekfeldt JDS, Cozzolino V, Kundel D, Kulhánek M, Mosimann C, et al. (2017). Potential of three microbial bio-effectors to promote maize growth and nutrient acquisition from alternative phosphorous fertilizers in contrasting soils. Chem Biol Technol Agri. 4:1–16.
- Ait-El-Mokhtar, M, Labbilta T, Anli M, Boutasknit A, Abouliatim Y, Khouloud M, et al. (2022c). Phosphate glass-based controlled-release fertilizers improve wheat growth, yield and grain nutritional quality under field conditions. Gesunde Pflanzen, 74:715–727.
- Mahmood M, Xu T, Ahmed W, Yang J, Li J, Mehmood S, et al. (2022). Variability in Soil Parent Materials at Different Development Stages Controlled Phosphorus Fractions and Its Uptake by Maize Crop. Sustainability. 14:5048.
- Reid TE, Kavamura VN, Abadie M, Torres-Ballesteros A, Pawlett M, Clark IM, et al. (2021). Inorganic Chemical Fertilizer Application to Wheat Reduces the Abundance of Putative Plant Growth-Promoting Rhizobacteria. Front Microbiol. 12:642587.
- Ben-Laouane R, Ait-El-Mokhtar M, Anli M, Boutasknit A, Ait Rahou Y, Raklami A, et al. (2021). Green compost combined with mycorrhizae and rhizobia: A strategy for improving alfalfa growth and yield under field conditions. Gesunde Pflanzen. 73:193–207.
- Ullah N, Ditta A, Imtiaz M, Li X, Jan AU, Mehmood S, Rizwan MS, Rizwan M. (2021). Appraisal for organic amendments and plant growth-promoting rhizobacteria to enhance crop productivity under drought stress: A review. J Agro and Crop Sci. 207:783–802.
- Cozzolino V, Monda H, Savy D, Di Meo V, Vinci G, Smalla K. (2021). Cooperation among phosphate-solubilizing bacteria, humic acids and arbuscular mycorrhizal fungi induces soil microbiome shifts and enhances plant nutrient uptake. Chem Biol Technol Agri. 8(1):1–18.
- Vahedi R, Rasouli-Sadaghiani M, Barin M, Vetukuri RR. (2021). Interactions between biochar and compost treatment and mycorrhizal fungi to improve the qualitative properties of a calcareous soil under rhizobox condition. Agriculture. 11:993.
- Huang HS, Chen QY, Liu LN, Cai P, Liang W, Li M. (2010). Poultry Manure Compost Alleviates the Phytotoxicity of Soil Cadmium: Influence on Growth of Pakchoi (Brassica chinensis L.). Pedosphere. 20(1):63–70.
- Mata-González R, Sosebee RE, Wan C. (2002). Shoot and root biomass of desert grasses as affected by biosolids application. J Arid Environ. 50:477–488.
- Schütz L, Gattinger A, Meier M, Müller A, Boller T, Mäder P, et al. (2018). Improving crop yield and nutrient use efficiency via biofertilization—A global meta-analysis. Front Plant Sci. 8:2204.
- Anli M, Symanczik S, El Abbassi A, Ait-El-Mokhtar M, Boutasknit A, Ben-Laouane R, et al. (2021). Use of arbuscular mycorrhizal fungus Rhizoglomus irregulare and compost to improve growth and physiological responses of Phoenix dactylifera ‘Boufgouss.’ Plant Biosystems. Int J Dealing Aspects Plant Biol. 155:763–771.
- Boutasknit A, Anli M, Tahiri A, Raklami A, Ait-El-Mokhtar M, Ben-Laouane R, et al. (2020a), Potential effect of horse manure-green waste and olive pomace-green waste composts on physiology and yield of garlic (Allium sativum L.) and soil fertility. Gesunde Pflanzen. 72:285–295.
- Chaichi W, Djazouli Z, Zebib B, Merah O (2018) Effect of vermicompost tea on faba bean growth and yield. Compost Sci Utilization. 26:279–285.
- Abd–Elrahman SH, Saudy HS, El–Fattah DAA, Hashem FA. (2022). Effect of irrigation water and organic fertilizer on reducing nitrate accumulation and boosting lettuce productivity. J Soil Sci Plant Nutr. 22:2144-2155.
- Ait-El-Mokhtar M, Fakhech A, Ben-Laouane R, Anli M, Boutasknit A, Ait-Rahou Y, Wahbi S, et al. (2022b). Compost as an eco-friendly alternative to mitigate salt-induced effects on growth, nutritional, physiological and biochemical responses of date palm. Document Type : Original. IJROWA. 11:85–100.
- Elfadil S, Meddich A, Boutasknit A, Anli M, Bouchdoug M, Jaouad A. (2022). Roles of various composts based on phosphate flotation waste, phosphogypsum and cactus in improving the tolerance of tomato plants to drought stress. J Mater Cycles Waste Manag. 24(5):1832-1841.
- Boutasknit A, Baslam M, Ait-El-Mokhtar M, Anli M, Ben-Laouane R, Douira A, et al. (2020b). Arbuscular mycorrhizal fungi mediate drought tolerance and recovery in two contrasting carob (Ceratonia siliqua L.) ecotypes by regulating stomatal, water relations, and (In)organic adjustments. Plants. 9:80.
- Cozzolino V, Di Meo V, Monda H, Spaccini R, Piccolo A. (2016). The molecular characteristics of compost affect plant growth, arbuscular mycorrhizal fungi, and soil microbial community composition. Biol Fertility Soils. 52 (1):15–29.
- Chouyia FE, Fiorentino N, Rouphael Y, Ventorino V, Fechtali T, Visconti D, et al. (2022). Assessing the effect of P-solubilizing bacteria and mycorrhizal fungi on tomato yield and quality under different crop rotations. Sci Horti. 293:110740.
- Lahbouki S, Ben-Laouane R, Anli M, Boutasknit A, Ait-Rahou Y, Ait-El-Mokhtar M, et al. (2022). Arbuscular mycorrhizal fungi and/or organic amendment enhance the tolerance of prickly pear (Opuntia ficus-indica) under drought stress. J Arid Environ. 199:104703.
- Zou YN, Srivastava AK, Ni QD, Wu QS. (2015). Disruption of mycorrhizal extraradical mycelium and changes in leaf water status and soil aggregate stability in rootbox-grown trifoliate orange. Front Microbiol. 6:203.
- .Mobaligh M, Meddich A, Imziln B, Fares K. (2021).The Use of Phosphate Washing Sludge to Recover by Composting the Leachate from the Controlled Landfill. Processes. 9(10):1735.
- McGonigle TP, Miller MH, Evans DG, Fairchild GL, Swan JA. (1990). A new method which gives an objective measure of colonization of roots by vesicular- arbuscular mycorrhizal Fungi. New Phytol. 115:495–501.
- The Application of Medicinal Plants in Traditional and Modern Medicine: A Review of Thymus vulgaris. Int J Clin Med. 6.
- Arnon DI. (1949). Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 24:1–15.
- Dhindsa RS, Plumbdhindsa P, Thorpe TA. (1981). Leaf senescence - Correlated with increased levels of membrane-permeability and lipid-peroxidation, and decreased levels of superoxide-dismutase and catalase. J Exp Bot. 32:93–101.
- Dubois M, Gilles KA, Hamilton JK, Rebers PT, Smith F. (1956). Colorimetric method for determination of sugars and related substances. Anal Chem. 28(3):350-356.
- Bradford MM. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 72:248–254.
- Polle A, Otter T, Seifert F. (1994). Apoplastic Peroxidases and Lignification in Needles of Norway Spruce (Picea abies L.). Plant Physiol. 1:53-60.
- Hori K, Wada ASTC. (1970). NII-Electronic Library Service. Chem Pharm Bull. 43:2091.
- Aebi H. (1984). Catalase in vitro. Methods Enzymol. 105:121–126.
- Nelson DW, Sommers LE. (1996). Total carbon, organic carbon, and organic matter. In: Methods of Soil Analysis, Part 3: Chemical Methods. Soil Sci. 961–1010
- Olsen S, Sommers L. (1982). Methods of soil analysis. Part 2. Chemical and microbiological properties of phosphorus. ASA Monograp. 9:403–430
- Khan I, Awan SA, Ikram R, Rizwan M, Akhtar N, Yasmin H, et al. (2021). Effects of 24-epibrassinolide on plant growth, antioxidants defense system, and endogenous hormones in two wheat varieties under drought stress. Physiologia Plantarum. 172(2):696–706.
- Köhl L, Lukasiewicz CE, van der Heijden MG. (2016). Establishment and effectiveness of inoculated arbuscular mycorrhizal fungi in agricultural soils. Plant Cell Envir. 39(1):136–146.
- van der Heijden MGA, Martin FM, Selosse MA, Sanders IR. (2015). Mycorrhizal ecology and evolution: the past, the present, and the future. New Phytol. 205:1406–1423.
- Hashem A, Kumar A, Al-Dbass AM, Alqarawi AA, Al-Arjani AF, Singh G, et al. (2019). Arbuscular mycorrhizal fungi and biochar improves drought tolerance in chickpea. Saudi J Biol Sci. 26 (3). 614–624.
- Meddich A, Jaiti F, Bourzik W, El Asli A, Hafidi M. (2015). Use of mycorrhizal fungi as a strategy for improving the drought tolerance in date palm (Phoenix dactylifera). Sci Horticulturae. 192:468–474.
- Zhang F, Zou YN, Wu QS. (2018). Quantitative estimation of water uptake by mycorrhizal extraradical hyphae in citrus under drought stress. Sci Horticulturae. 229:132–136.
- Wu QS, Srivastava AK, Zou YN. (2013). AMF-induced tolerance to drought stress in citrus: A review. Scientia Horticulturae. 164:77–87.
- Kilpeläinen J, Aphalo PJ, Lehto T. (2020). Temperature affected the formation of arbuscular mycorrhizas and ectomycorrhizas in Populus angustifolia seedlings more than a mild drought. Soil Biol Biochem:146.
- Anli M, Boutasknit A, Ait-El-Mokhtar M, Ben-Laouane R, Ait-Rahou Y, Fakhech A, et al. (2022). Improving lettuce yield and quality of an agricultural soil using a combination of arbuscular mycorrhizal fungus and phosphate-green wastes compost. Gesunde Pflanzen. 74:205–217.
- Ziane H, Hamza N, Meddad-Hamza A. (2021). Arbuscular mycorrhizal fungi and fertilization rates optimize tomato (Solanum lycopersicum L.) growth and yield in a Mediterranean agroecosystem. J Saudi Society Agri Sci. 20:454–458.
- Pinos NQ, Louro Berbara RL, Elias SS, van Tol de Castro TA, García AC. (2019). Combination of humic substances and arbuscular mycorrhizal fungi affecting corn plant growth. J Enviro Quality. 48 (6):1594–1604.
- Verma P, Yadav AN, Khannam KS, Panjiar N, Kumar S, Saxena AK, Suman A. (2015). Assessment of genetic diversity and plant growth promoting attributes of psychrotolerant bacteria allied with wheat (Triticum aestivum) from the northern hills zone of India. Annals Microbiol. 65:1885–1899.
- Benaffari W, Boutasknit A, Anli M, Ait-El-Mokhtar M, Ait-Rahou Y, Ben-Laouane R, et al. (2022). The native arbuscular mycorrhizal fungi and vermicompost-based organic amendments enhance soil fertility, growth performance, and the drought stress tolerance of quinoa. Plants. 11:393.
- Anli M, Baslam M, Tahiri A, Raklami A, Symanczik S, Boutasknit A, et al (2020). Biofertilizers as strategies to improve photosynthetic apparatus, growth, and drought stress tolerance in the date palm. Front Plant Sci. 11:516818.
- Mandal D, Pal R. (2022). Ecology of rhizosphere mycoflora in organic matter amended soil : A review. Pharma Innov J. 11:3677–3679.
- Olayiwola VA, Abiodun FO. (2019). Effects of Arbuscular Mycorrhiza Fungi (Glomus mossae) and Compost on Early Growth Performance of Parkia biglobosa. Curr J Appl Sci Technol. 37(4):1–8.
- Duo LA, Liu CX, Zhao SL. (2018). Alleviation of drought stress in turfgrass by the combined application of nano-compost and microbes from compost. Russian J Plant Physiol. 65(3):419–426.
- Ortuño MF, Lorente B, Hernández JA, Sánchez-Blanco MJ. (2018). Mycorrhizal inoculation on compost substrate affects nutritional balance, water uptake and photosynthetic efficiency in Cistus albidus plants submitted to water stress. Rev Brasileira Botanica. 41(2):299–310.
- Boutasknit A, Baslam M, Ait-El-Mokhtar M, Anli M, Ben-Laouane R, Ait-Rahou Y, et al. (2021). Assemblage of indigenous arbuscular mycorrhizal fungi and green waste compost enhance drought stress tolerance in carob (Ceratonia siliqua L.) trees. Sci Rep. 11:22835.
- Symanczik S, Lehmann MF, Wiemken A, Boller T, Courty PE. (2018). Effects of two contrasted arbuscular mycorrhizal fungal isolates on nutrient uptake by Sorghum bicolor under drought. Mycorrhiza. 28:779–785.
- Lovisolo C, Perrone I, Carra A, Ferrandino A, Flexas J, Medrano H, et al. (2010). Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: A physiological and molecular update. Functional Plant Biol. 37(2):98–116.
- Bouchemal K, Bouldjadj R, Belbekri MN, Ykhlef N, Djekoun A. (2018). Photosynthetic pigments, antioxydant enzymes and leaf osmotic potential of ten durum wheat (Triticum durum). genotypes: impact of water stress. Phytoprotection 98:13–24.
- Morales F, Ancín M, Fakhet D, González-Torralba J, Gámez AL, Seminario A, et al. (2020). Photosynthetic Metabolism under Stressful Growth Conditions as a Bases for Crop Breeding and Yield Improvement. Plants (Basel). 9(1):88.
- Yang X, Li Y, Chen H, Huang J, Zhang Y, Qi M, et al. (2020). Photosynthetic Response Mechanism of Soil Salinity-Induced Cross-Tolerance to Subsequent Drought Stress in Tomato Plants. Plants. 9:363.
- Shafiq S, Akram NA, Ashraf M, García-Caparrós P, Ali OM, Latef AAHA. (2021). Influence of Glycine Betaine (Natural and Synthetic) on Growth, Metabolism and Yield Production of Drought-Stressed Maize (Zea mays L.) Plants. Plants. 10:2540.
- Al-Arjani ABF, Hashem A, Abd_Allah EF. (2020). Arbuscular mycorrhizal fungi modulates dynamics tolerance expression to mitigate drought stress in Ephedra foliata Boiss. Saudi J Biol Sci. 27:380–394.
- Hasanuzzaman M, Bhuyan MHMB, Zulfiqar F, Raza A, Mohsin SM, Mahmud JA, et al. (2020). Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants. 9(8):681.
- Kiani SP, Maury P, Sarrafi A, Grieu P. (2008). QTL analysis of chlorophyll fluorescence parameters in sunflower (Helianthus annuus L.) under well-watered and water-stressed conditions. Plant Sci. 175(4):565–573.
- Tian F, Wang W, Liang C, Wang X, Wang G, Wang W. (2017). Overaccumulation of glycine betaine makes the function of the thylakoid membrane better in wheat under salt stress. Crop J. 5:73–82.
- Bárzana G, Aroca R, Bienert GP, Chaumont F, Ruiz-Lozano JM. (2014). New insights into the regulation of aquaporins by the arbuscular mycorrhizal symbiosis in maize plants under drought stress and possible implications for plant performance. Mol Plant Microbe Interact. 27:349–363.
- Muhammad F, Romdhane L, Al Sulti MKRA, Rehman A, Al-Busaidi WM, Lee DJ. (2020). Morphological, physiological and biochemical aspects of osmopriming-induced drought tolerance in lentil. J Agro Crop Sci. 206(2):176–186.
- Thalmann M, Santelia D. (2017). Starch as a determinant of plant fitness under abiotic stress. New Phytologist. 214:943–951.
- Sairam RK, Rao K, Srivastava GC. (2002). Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 163(5):1037–1046.
- Huang YM, Srivastava AK, Zou YN, Ni QD, Han Y, Wu QS. (2014). Mycorrhizal-induced calmodulin mediated changes in antioxidant enzymes and growth response of drought-stressed trifoliate orange. Frontiers Microbiol. 5:1–7.
- Ni QD, Zou YN, Wu QS, Huang YM. (2013). Increased tolerance of citrus (Citrus Tangerina) seedlings to soil water deficit after mycorrhizal inoculation: Changes in antioxidant enzyme defense system. Notulae Botanicae Horti Agrobotanici Cluj-Napoca. 41(2):524–529.
- Rani B, Madan S, Sharma KD, Pooja Kumar A. (2018). Influence of arbuscular mycorrhiza on antioxidative system of wheat (Triticum aestivum) under drought stress. Ind J Agri Sci. 88 (2):289–295.
- Ruiz-Lozano JM. (2003). Arbuscular mycorrhizal symbiosis and alleviation of osmotic stress. New perspectives for molecular studies. Mycorrhiza. 13(6):309–317.
- Tahiri A, Meddich A, Raklami A, Alahmad A, Bechtaoui N, Anli M, et al. (2022). Assessing the potential role of compost, PGPR, and AMF in improving tomato plant growth, yield, fruit quality, and water stress tolerance. J Soil Sci Plant Nutr. 22:743–764.
- Yadav SK, Dhote M, Kumar P, Sharma J, Chakrabarti T, Juwarkar AA. (2010). Differential antioxidative enzyme responses of Jatropha curcas L. to chromium stress. J Hazard Mater. 180:609–615.
- Hasanuzzaman M, Parvin K, Bardhan K, Nahar K, Anee TI, Masud AAC, et al. (2021). Biostimulants for the regulation of reactive oxygen species metabolism in plants under abiotic stress. Cells. 10 (10):1–29.
- Djébali N, Mhadhbi H, Lafitte C, Dumas B, Esquerré-Tugayé MT, Aouani ME, et al. (2011). Hydrogen peroxide scavenging mechanisms are components of Medicago truncatula partial resistance to Aphanomyces euteiches. Eur J Plant Pathol. 131(4):559–571.
- Du B, Kruse J, Winkler JB, Alfarray S, Schnitzler J, Ache P, et al. (2019). Climate and development modulate the metabolome and antioxidative system of date palm leaves. J Exp Bot. 70(20):5959–5969.
- Ahmad P. (2014). Oxidative damage to plants: antioxidant networks and signaling.
- Kaushal M. (2019). Microbes in cahoots with plants: MIST to hit the jackpot of agricultural productivity during drought. Int J Mol Sci. 20 (7):1769.
- Wang JY, Lin PY, Al-Babili S. (2021). On the biosynthesis and evolution of apocarotenoid plant growth regulators. Semin Cell Dev Biol. 109:3–11.
- Kevin O, Joyce JL, Samuel MM. (2021). Soil properties and maize (Zea mays L.) growth and yield response to water hyacinth (Eichhornia crassipes) compost application in Lake Victoria Basin, Kenya. J Soil Sci Envir Manag. 12(1):17–28.
- Sun J, Zou L, Li W, Yang J, Wang Y, Xia Q, et al. (2018). Rhizosphere soil properties and banana Fusarium wilt suppression influenced by combined chemical and organic fertilizations. Agri Ecosystems Envir. 254:60–68.
- Yu Y, Wang C, Shi Y, Li M, Zhang L, Sun Z, et al. (2022). Land-Use Type Affects Soil Microbial Community Assembly Process and Inter-Kingdom Co-Occurrence Pattern in a Floodplain Ecosystem. SSRN Electronic J. 179:104574.